Describe C4 pathway of photosynthesis. How is this pathway an adaptive advantage to the plant ?

C4 carbon fixation is one of three biochemical mechanisms, along with C3 and CAM photosynthesis, used in carbon fixation. It is named for the 4-carbon molecule present in the first product of carbon fixation in the small subset of plants known as C4 plants, in contrast to the 3-carbon molecule products in C3 plants.
C4 fixation is an elaboration of the more common C3 carbon fixation and is believed to have evolved more recently. C4 and CAM overcome the tendency of the enzyme RuBisCO to wastefully fix oxygen rather than carbon dioxide in what is called photorespiration. This is achieved by using a more efficient enzyme to fix CO2 in mesophyll cells and shuttling this fixed carbon via malate or aspartate to bundle-sheath cells. In these bundle-sheath cells, RuBisCO is isolated from atmospheric oxygen and saturated with the CO2 released by decarboxylation of the malate or oxaloacetate. These additional steps, however, require more energy in the form of ATP. Because of this extra energy requirement, C4 plants are able to more efficiently fix carbon in only certain conditions, with the more common C3 pathway being more efficient in other conditions.
C4 pathwayThe first experiments indicating that some plants do not use the established C3 carbon fixation but produce malate and aspartate in the first step were done in the 1950s and early 1960s by Hugo P. Kortschak[1] and Yuri Karpilov[2] The C4 pathway was finally discovered by Marshall Davidson Hatch and C. R. Slack, in Australia, in 1966, so it is sometimes called the Hatch-Slack pathway.[3]
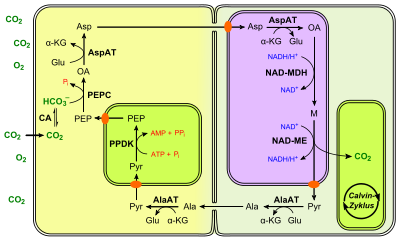
In C3 plants, the first step in the light-independent reactions of photosynthesis involves the fixation of CO2 by the enzyme RuBisCO into 3-phosphoglycerate. However, due to the dual carboxylase and oxygenase activity of RuBisCo, an amount of the substrate is oxidized rather than carboxylated, resulting in loss of substrate and consumption of energy, in what is known as photorespiration. In order to bypass the photorespiration pathway, C4 plants have developed a mechanism to efficiently deliver CO2 to the RuBisCO enzyme. They utilize their specific leaf anatomy where chloroplasts exist not only in the mesophyll cells in the outer part of their leaves but in the bundle sheath cells as well. Instead of direct fixation to RuBisCO in the Calvin cycle, CO2 is incorporated into a 4-carbon organic acid, which has the ability to regenerate CO2 in the chloroplasts of the bundle sheath cells. Bundle sheath cells can then utilize this CO2 to generate carbohydrates by the conventional C3 pathway.
The first step in the pathway is the conversion of pyruvate to phosphoenolpyruvate (PEP), by the enzyme pyruvate orthophosphate dikinase. This reaction requires inorganic phosphate and ATP plus pyruvate, producing phosphoenolpyruvate, AMP, and inorganic pyrophosphate (PPi). The next step is the fixation of CO2 into oxaloacetate by the enzyme PEP carboxylase. Both of these steps occur in the mesophyll cells:
pyruvate + Pi + ATP → PEP + AMP + PPiPEP + CO2 → oxaloacetatePEP carboxylase has a lower Km for CO2 — and, hence, higher affinity — than RuBisCO. Furthermore, O2 is a very poor substrate for this enzyme. Thus, at relatively low concentrations of CO2, most CO2 will be fixed by this pathway.
The product is usually converted to malate, a simple organic compound, which is transported to the bundle-sheath cells surrounding a nearby vein. Here, it is decarboxylated to produce CO2 and pyruvate. The CO2 now enters the Calvin cycle and the pyruvate is transported back to the mesophyll cell.
Since every CO2 molecule has to be fixed twice, first by 4-carbon organic acid and second by RuBisCO, the C4 pathway uses more energy than the C3 pathway. The C3 pathway requires 18 molecules of ATP for the synthesis of one molecule of glucose, whereas the C4 pathway requires 30 molecules of ATP. This energy debt is more than paid for by avoiding losing more than half of photosynthetic carbon in photorespiration as occurs in some tropical plants,[citation needed] making it an adaptive mechanism for minimizing the loss.
There are several variants of this pathway:
- The 4-carbon acid transported from mesophyll cells may be malate, as above, or aspartate
- The 3-carbon acid transported back from bundle-sheath cells may be pyruvate, as above, or alanine
- The enzyme that catalyses decarboxylation in bundle-sheath cells differs. In maize and sugarcane, the enzyme is NADP-malic enzyme; in millet, it is NAD-malic enzyme; and, in Panicum maximum, it is PEP carboxykinase